27.3. The influence of vocal folds stiffness on the modelled
EGG waveform
In the two-mass model control
of the fundamental frequency can be exercised in two ways. The first approach
is to add a "tension parameter" Q (Ishizaka & Flanagan
1972:1279), the second is to correct the vocal folds stiffness directly and
according to muscular action.
In the first method,
the mass and thickness of the folds are scaled down by the factor Q,
whereas the tensions k are scaled up, "causing the fundamental
frequency to vary proportionally with Q. Phase difference, duty ratio, and
the glottal area waveforms are essentially uninfluenced by Q, and the amplitudes
of glottal area and glottal flow decrease gradually with increasing Q"
(Ishizaka & Flanagan, 1972:1259). As is known from several
experiments the Open Quotient increases with the increase in fundamental
frequency. This effect has been included in the model by modification of
the coupling tension kc. The kc variation was empirically
determined to be proportional to Q2. The Q factor is related
to the stretching of the vocal folds (by contraction of the cricothyroid
muscle) and contracting the vocalis muscle (see section 27.3 for details).
The modifications of the model which are closely related to Q are
in accordance with the experimental measuremnts: "Variation of the duty
ratio with frequency falls into the range measured in inverse filtering
experiments" (Ishizaka and Flanagan, 1972:1260).
The simulation of the EGG waveform for a varying
Q factor was performed for neutral /e:/ with Q varying
from 0.8 to 1.4 in 0.1 steps and other parameters remaining unchanged (exactly
as in section 27.1). In particular, the subglottal pressure Ps was
set to 8 cm H2O. The increase in the Q factor leads to an increase in
F0 from 115 to 190 Hz. The simulated EGG waveforms exhibit the expected
dependencies and correlate well with the modelled glottal airflow.
-
Figure 52. The dependency of the duty ratio of the
modelled glottal pulse on the Q factor: a) measured in the glottal
volume velocity Ug waveform, b) measured in the EGG waveform as
OQI c) and d) measured as OQII in the glottal volume velocity
Ug waveform and in the EGG waveform, respectively.
|

 |
The relations between the glottal flow Open Quotient
and the Open Quotient of the EGG are depicted in Fig.
52. The vocal folds open longer
when the muscular tension increases. The same tendency can be
observed in the EGG waveform, however, the results by the second measure
of the Open Quotient (section 12) are strongly related to the opening and
closing angles of the folds. If the closing angle
thetac and the opening angle thetao are
relatively (very relatively) wide (thetac > 1º
and thetao > 2.5º), a complementary effect is observed,
i.e. the EGG duty ratio (especially OQII) increases with increasing
muscular tension. This should not be overestimated; it is to be expected
that higher muscular tension causes rather narrower angles of the folds opening
and closing. However, the appropriate physiological measurements are not
given in the literature.
The skewness of the modelled glottal airflow pulse
decreases with increased pitch as can be see in Fig.
50a and Fig.53a.
This is in accordance with the results of the glottal modelling of Ishizaka
and Flanagan (1972), the body-cover models of Titze (1992, 1994) and the
inverse filtering measurements of Koreman (1996:157-159), but contradictory
to the results of Pierrehumbert's inverse filtering (1989) and Sluijter's
speech signal measurements (1995:125). Pierrehumbert (1989) explained the
difference between the experimental outcome and the two-mass model with the
increase in the coupling stiffness kc between the vibrating masses,
which in turn is related to the tension in the vocalis muscle (thyroarytenoid
muscle). The experiments conducted with the two-mass model do not confirm
this explanation. In the two-mass model the stiffness kc changes
with Q2 and the skewness of the glottal pulse decreases with the
increase of F0 and OQ. A better explanation for the effect
is the simultaneous action of two sources: the contraction of muscles and
the increased subglottal pressure. Higher muscular tension
effects the increase in the glottal airflow duty ratio, while the higher
skewness of the glottal pulse is caused by an increased subglottal
pressure. The increase of the subglottal pressure has already
been correlated with the linguistic marking of word stress, while muscular
tension has been anticipated to correlate with the prosodic function of sentence
intonation. This hypothesis is in accordance with Pierrehumbert's results
(1989), because the examined intonation contours (F0 changes) are superimposed
on the word stress.
-
Figure 53. Dependency of the skewness of the modelled
glottal pulse on the Q tension factor: a) Speed Quotient measured
in the glottal volume velocity Ug waveform, b) steepness of the
contact rise and fall measured in the EGG waveform
|

 |
Other results of the EGG modelling support this hypothesis,
too. As shown in Fig. 54b, the steepness of the contact
rise and fall does not depend on the increased tension of the vocal folds.
This is in agreement with the outcome our experiments (chapter III), where
the influence of intonation patterns on vowels in stressed positions was
measured. This allows us to formulate the thesis
that the increase in subglottal pressure is positively correlated with the
steepness of the EGG pulse flanks, whereas the rise of the duty ratio of
the EGG waveform depends rather on the increase in muscular
tension (increased stiffness of the vocal folds). Both mechanisms
also cause a rise of the fundamental frequency.
The second type of vocal folds tension modification
follows the body-cover model (Titze, 1994) and provides a more detailed
view of the physiology of fundamental frequency control. Pierrhumbert's (1989)
remark that "in the mid to high region of the pitch range, the dominant
mechanism for F0 control appears to be cricothyroid contraction
and relaxation" can be applied to the model. Generally, the stiffness
of the vocal folds is controlled mainly by the actions of the cricothyroid
and thyroarytenoid muscles (Titze, 1994:202-209). The theory of the
F0 control mechanism introduced by Titze (1988, 1994) explains
F0 variations in terms of the body-cover model of the vocal folds.
The F0 changes in modal phonation with normal voice effort are primarily
caused by the action of the thyroarytenoid muscle. The muscle can stiffen
the body of the vocal folds while slackening the cover. That is why the
F0 changes depend on the type of vibrating tissue, because the vibrating
tissue is primarily muscular in the cross-section (the body). It is to be
expected that the effective stiffness of the vibrating mass will rise due
to the thyroarytenoid contraction. If, however, the non-muscular part of
the folds (the cover) vibrates, then the effective stiffness will presumably
be lowered because of the contraction of the thyroarytenoid muscle (Titze,
1994:206). This causes positive or negative changes in F0 depending
on the type of vibrating tissue. Titze (1994:206) relates the
changes in the fundamental frequency to the relative
thickness of that part of the muscular layer which is vibrating:

where da/d is the ratio of the depth
of the TA muscle in vibration to the total depth in vibration,
sigmaam denotes the maximum possible stress of the
thyroarytenoid, sigmap is the effective passive stress
of all tissue in vibration (the stress during the total inactivity of the
thyroarytenoid). Lm is the length of the mucous part of the folds,
rho is the tissue density (1.04 kg/m3) and aTA
represents the activity of the thyroarytenoid muscle. Titze (1994:208) gives
the average dependency of F0 on muscle activity and notes that in
speech production "cricothyroid and thyroarytenoid activity are both
relatively low (<50 percent of maximum), and changes in F0 with
increased thyroarytenoid activity are all positive, in agreement with average
correlations". It can be concluded that in the
normal speech phonation modus, the rise in F0 results from higher
thyroarytenoid activity while cricothyroid activity is low.
Muscle activities and their influence on fundamental
frequency are usually modelled in the continuous model (Berry & Titze,
1996; Titze & Talkin, 1979) or the 3-mass model (Story & Titze, 1995).
However, Lucero (1993, 1996) shows that the different mechanical properties
of body and cover can be modelled even using the standard configuration of
the two-mass model. Taking his simplification into
consideration, the stiffness of the vibrating masses m1 and
m2 then depends on the contraction of the cricothyroid and
thyroarytenoid muscles, in the sense that their action can be regarded as
additive in the lower mass and contrary (differential) in the upper mass.
Stiffness is then expressed as k1=CT+TA and
k2=CT-TA, where CT and TA represent the
weighted activities of the cricothyroid and thyroarytenoid muscles
(Lucero, 1996). During modal phonation, TA is relatively high
and CT relatively low, which results in high values for k1 and low
values for k2. k1 resembles the kb parameter of
the body-cover model, in that "increasing of the body stiffness (i.e.
contracting the TA) leads to lower body layer amplitude and higher
pitches" (Story & Titze, 1996:1256).
These relations are also included
in our model, as the influence of muscular tension on the simulated EGG waveform
is investigated. As before, the model settings are kept constant,
while only the tension parameters k1, k2 and kc
are modified. Neutral [ ] is modelled and the parameters
are set to the values given in section 27.1. In particular, the opening and
closing angles were set to thetao = 1.0º and
thetac = 0.5º.
] is modelled and the parameters
are set to the values given in section 27.1. In particular, the opening and
closing angles were set to thetao = 1.0º and
thetac = 0.5º.
The tensions are
set to the following values:
| k1 (kdyn) |
k2 (dyn) |
kc (kdyn) |
0.8 |
800 |
55 |
8 |
40 |
55 |
120 |
4 |
55 |
220 |
0.4 |
55 |
300 |
0.2 |
55 |
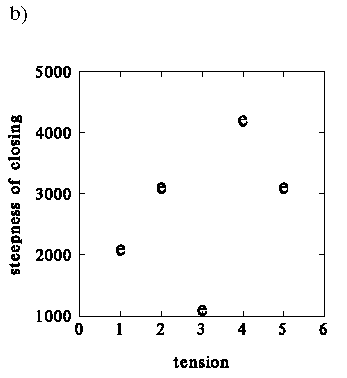
The results of the simulation are depicted in Fig.
54a and b. Generally, the results
are similar to those produced by Q factor modelling.
The simulated increased tension of the cricothyroid and thyroarytenoid muscles
causes the increase of the Open Quotient in both Ug and EGG waveforms.
The steepness of the EGG slopes remains almost unchanged, whereas the skewness
of the glottal flow decreases.
-
Figure 54. Dependency of the duty ratio and the steepness
of the modelled EGG pulse on the tension of the vocal folds: a) Open Quotient
of the EGG computed using method II b) steepness of the contact rise measured
in the EGG waveform
|

 |
Titze (1990) has formulated the dependencies of the contact area on the motion
of the folds6.
Compared to the nominal (typical) vocal folds
configuration, the simulated EGG pulses are
-
wider, if there is a uniform displacement of the
vocal fold towards the midline,
-
skewed to the left, if the glottis is more convergent
and there is vertical phasing of the folds occurs,
-
more rectangular (steeper slopes and longer, gradual
changes in the maximum contact phase), if one of the folds is bulging,
-
more triangular, if there is vertical phasing of
the fold (a large phase difference between lower and upper margin makes the
pulse more triangular. A small phase angle, on the other hand, makes the
waveform more rectangular).
6 Titze's conclusions are
based on the continuous model of vocal folds motion.